Bipedalism is marked by a number of anatomical modifications to the standard primate body. These center on the pelvis and involve changes in the head (cranium) and the rest of the body (postcranium), reflecting a shifting of the center of balance from the abdominal cavity to the hip. In mammals, the hip is composed of three mirrored sets of bones: the ilium, the ischium and the pubis (Figure 1). The top part of the leg fits into the bottom-rear portion of the ilium, into a round socket called the acetabulum. It is one of two ball-and-socket joints in the body, the other being where the arm fits into the scapula at the top of your shoulder.
Bipedalism is marked by a number of anatomical modifications to the standard primate body. These center on the pelvis and involve changes in the head (cranium) and the rest of the body (postcranium), reflecting a shifting of the center of balance from the abdominal cavity to the hip. In mammals, the hip is composed of three mirrored sets of bones: the ilium, the ischium and the pubis (Figure 1). The top part of the leg fits into the bottom-rear portion of the ilium, into a round socket called the acetabulum. It is one of two ball-and-socket joints in the body, the other being where the arm fits into the scapula at the top of your shoulder. Other changes are present above the hip, as well. Because they are quadrupedal, chimpanzees, gorillas and orangutans have a straight backbone, or vertebral column (Figure 3). In humans, the vertebral column resembles a double “s” shape, which balances the torso above the hip (and creates the back problems we suffer later in life). At the top of the spinal column, the top vertebra, the atlas, has facets that balance the head and the second vertebra, the axis, has a prong that fits directly into a hole in the skull. This hole, which is called the foramen magnum, is at the back of the skull in higher apes, as in all quadrupedal animals. This allows the animal to keep its head up while it is trotting along the ground. In humans, the foramen magnum is at the base of the skull, allowing us to look forward as we walk. It also makes it hard to look forward when we crawl on all fours. Each of these modifications is diagnostic of humans and easily recognizable in the fossil record in specimens for which these anatomical areas are present.
Other changes are present above the hip, as well. Because they are quadrupedal, chimpanzees, gorillas and orangutans have a straight backbone, or vertebral column (Figure 3). In humans, the vertebral column resembles a double “s” shape, which balances the torso above the hip (and creates the back problems we suffer later in life). At the top of the spinal column, the top vertebra, the atlas, has facets that balance the head and the second vertebra, the axis, has a prong that fits directly into a hole in the skull. This hole, which is called the foramen magnum, is at the back of the skull in higher apes, as in all quadrupedal animals. This allows the animal to keep its head up while it is trotting along the ground. In humans, the foramen magnum is at the base of the skull, allowing us to look forward as we walk. It also makes it hard to look forward when we crawl on all fours. Each of these modifications is diagnostic of humans and easily recognizable in the fossil record in specimens for which these anatomical areas are present. In 1994, the remains of a remarkable hominin, dated to 4.4 million years ago, were unearthed in the Afar Triangle of Northeastern Ethiopia. Examination of the surrounding deposits, however, yielded a conclusion that this hominin lived in a woodland environment, rather than a savannah/forest fringe environment (White et al. 2009a). Requiring over ten years of extrication from the surrounding rock and painstaking reconstruction, this fossil form, Ardipithecus ramidus (now represented by 110 individuals) yields diagnostic parts of the pelvis (Figure 4), as well as sections of the arms and skull (Figure 5) (White et al. 2009b). Although the base of the skull is not preserved, one striking aspect of humanity is present in the teeth. The canine (eye tooth) does not extend beyond the tooth row. Humans are the only hominins for which this is the case. In all other ape species, fossil and extant, the canine projects well beyond the tooth row.
In 1994, the remains of a remarkable hominin, dated to 4.4 million years ago, were unearthed in the Afar Triangle of Northeastern Ethiopia. Examination of the surrounding deposits, however, yielded a conclusion that this hominin lived in a woodland environment, rather than a savannah/forest fringe environment (White et al. 2009a). Requiring over ten years of extrication from the surrounding rock and painstaking reconstruction, this fossil form, Ardipithecus ramidus (now represented by 110 individuals) yields diagnostic parts of the pelvis (Figure 4), as well as sections of the arms and skull (Figure 5) (White et al. 2009b). Although the base of the skull is not preserved, one striking aspect of humanity is present in the teeth. The canine (eye tooth) does not extend beyond the tooth row. Humans are the only hominins for which this is the case. In all other ape species, fossil and extant, the canine projects well beyond the tooth row.
 In the early 1920s, a young anatomist named Raymond Dart took a job at the University of Witwatersrand in Johannesburg, South Africa. Keenly interested in comparative primate anatomy, Dart had been advised to go to the Wit by the famed anatomist Sir Grafton Eliot Smith and, upon arrival, began work on the ancestry of South African primates.
In the early 1920s, a young anatomist named Raymond Dart took a job at the University of Witwatersrand in Johannesburg, South Africa. Keenly interested in comparative primate anatomy, Dart had been advised to go to the Wit by the famed anatomist Sir Grafton Eliot Smith and, upon arrival, began work on the ancestry of South African primates.

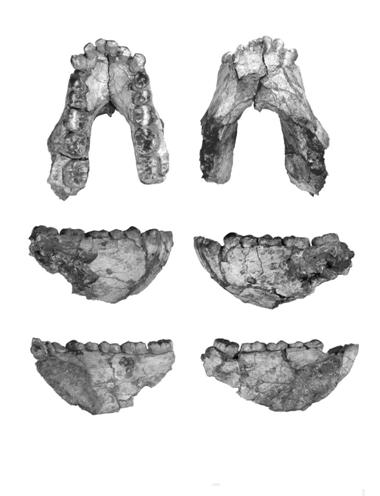

 In 1973, working with a local team of fossil hunters, Maurice Taieb went to an arid stretch of land in the Afar Triangle of Ethiopia to an area called Hadar. A year later, Don Johanson, a member of his team discovered one of the most famous of all fossil hominin discoveries ever made. Exploring 3.4 and 3.6 million years old deposits, he discovered the fragmentary remains that constituted 40% of the skeleton of a small adult female (Figure 4) . This individual was nicknamed “Lucy” after the Beatles’ song
In 1973, working with a local team of fossil hunters, Maurice Taieb went to an arid stretch of land in the Afar Triangle of Ethiopia to an area called Hadar. A year later, Don Johanson, a member of his team discovered one of the most famous of all fossil hominin discoveries ever made. Exploring 3.4 and 3.6 million years old deposits, he discovered the fragmentary remains that constituted 40% of the skeleton of a small adult female (Figure 4) . This individual was nicknamed “Lucy” after the Beatles’ song  When Lucy was examined, it was found that the shape and position of the the teeth and jaw as well as the hip and long bone fragments put her almost perfectly intermediate be tween the ape position and the human position. Although she had the overall size and rib cage structure of a chimpanzee, her pelvis and leg bones were perfectly adapted for bipedalism. It was in the teeth and palate that the clearest transitional characteristics existed (Figure 5, right). In modern humans (5c), the dental arcade (tooth row) is in the shape of a parabola, like the Gateway Arch in Saint Louis. In apes (5a), it is a sharp “U” shape. In A. afarensis (5b), it is intermediate, tending toward the ape condition.
When Lucy was examined, it was found that the shape and position of the the teeth and jaw as well as the hip and long bone fragments put her almost perfectly intermediate be tween the ape position and the human position. Although she had the overall size and rib cage structure of a chimpanzee, her pelvis and leg bones were perfectly adapted for bipedalism. It was in the teeth and palate that the clearest transitional characteristics existed (Figure 5, right). In modern humans (5c), the dental arcade (tooth row) is in the shape of a parabola, like the Gateway Arch in Saint Louis. In apes (5a), it is a sharp “U” shape. In A. afarensis (5b), it is intermediate, tending toward the ape condition. The case for habitual bipedalism received added support from a site much further south than where Lucy had been found. In 1976, Andrew Hill, a digger in Mary Leakey’s team, working at the site of Laetoli, in Tanzania, unearthed a set of hominin tracks that had been covered by a now extinct volcano. These footprints, which extend approximately 80 feet across the plain, have been securely dated at 3.6 million years and show where two individuals walked side-by-side (Figure 6). The tracks are significant in that they demonstrate that the individuals who made them had arches and practiced the characteristic “toe-off” pattern of gait practiced only by hominins. The presence of A. afarensis skeletal remains nearby at the same level provided the link to the footprints.
The case for habitual bipedalism received added support from a site much further south than where Lucy had been found. In 1976, Andrew Hill, a digger in Mary Leakey’s team, working at the site of Laetoli, in Tanzania, unearthed a set of hominin tracks that had been covered by a now extinct volcano. These footprints, which extend approximately 80 feet across the plain, have been securely dated at 3.6 million years and show where two individuals walked side-by-side (Figure 6). The tracks are significant in that they demonstrate that the individuals who made them had arches and practiced the characteristic “toe-off” pattern of gait practiced only by hominins. The presence of A. afarensis skeletal remains nearby at the same level provided the link to the footprints.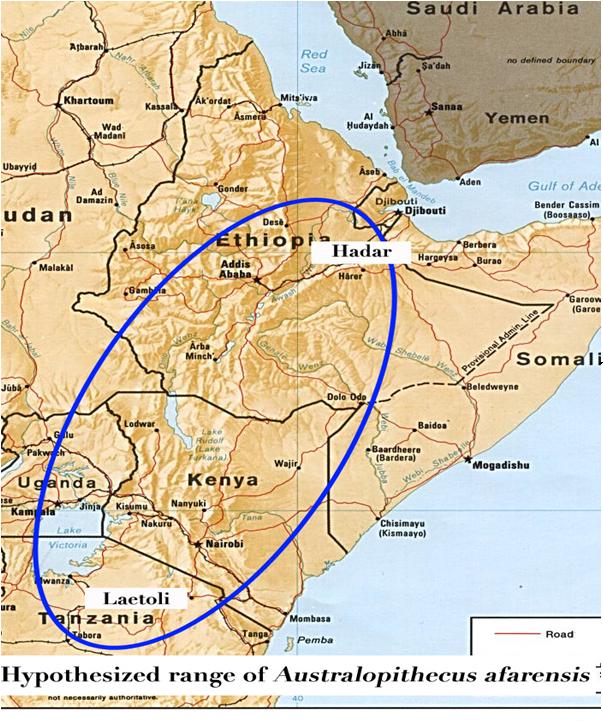

 The Piltdown forgery ranks as one of the best scientific hoaxes of all time. Charles Dawson unearthed purported hominin fossil remains from a gravel pit at Piltdown Commons, East Sussex County, in 1912 that consisted of a mostly complete skull and partial jaw in association with extinct mastodon and hippopotamus fossils (See Figure 1). This was published in the Quarterly Journal of the Geological Society (Dawson and Woodward 1913) and became the crown of English anthropology, eliciting the support of most of the top anthropologists and anatomists of the day, including Sir Arthur Keith, Sir Arthur Smith-Woodward, Grafton Eliot Smith and William King Gregory.
The Piltdown forgery ranks as one of the best scientific hoaxes of all time. Charles Dawson unearthed purported hominin fossil remains from a gravel pit at Piltdown Commons, East Sussex County, in 1912 that consisted of a mostly complete skull and partial jaw in association with extinct mastodon and hippopotamus fossils (See Figure 1). This was published in the Quarterly Journal of the Geological Society (Dawson and Woodward 1913) and became the crown of English anthropology, eliciting the support of most of the top anthropologists and anatomists of the day, including Sir Arthur Keith, Sir Arthur Smith-Woodward, Grafton Eliot Smith and William King Gregory.
 At around 2.5 million years ago, a new form was found in Ethiopia that appeared to be a scaled up version of A. afarensis. Called Australopithecus aethiopicus, this species had a long, low cranium with flared cheekbones and attachment areas for very powerful chewing muscles on its face (See Figure 3). To go with this, A. aethiopicus also had, on the top of its head, a large sagittal crest. Work with modern-day gorillas, which also have this feature, has shown that this is not a genetic trait but appears as a result of bone deposition on the top of the head through continuous grinding of nut and plant substances. Unlike the gorilla version, however, which is focused directly on the top of the cranium (vertex), the A. aethiopicus manifestation is toward the back of the cranium. This form had an average cranial capacity of around 410 cubic centimeters. For comparison purposes, the average modern human cranium is approximately 1450 cubic centimeters in capacity while chimpanzees average 375 cubic centimeters.
At around 2.5 million years ago, a new form was found in Ethiopia that appeared to be a scaled up version of A. afarensis. Called Australopithecus aethiopicus, this species had a long, low cranium with flared cheekbones and attachment areas for very powerful chewing muscles on its face (See Figure 3). To go with this, A. aethiopicus also had, on the top of its head, a large sagittal crest. Work with modern-day gorillas, which also have this feature, has shown that this is not a genetic trait but appears as a result of bone deposition on the top of the head through continuous grinding of nut and plant substances. Unlike the gorilla version, however, which is focused directly on the top of the cranium (vertex), the A. aethiopicus manifestation is toward the back of the cranium. This form had an average cranial capacity of around 410 cubic centimeters. For comparison purposes, the average modern human cranium is approximately 1450 cubic centimeters in capacity while chimpanzees average 375 cubic centimeters. Also from this time period is a form similar to A. afarensis in brain size and gracility, Australopithecus garhi (“surprise” in the Afar language). The teeth are slightly larger than those of afarensis (See Figure 4). In overall body shape, however A. garhi was far more modern, with the ratio of upper arm length to upper leg length much closer to those of later hominins (Asfaw et al. 1999).
Also from this time period is a form similar to A. afarensis in brain size and gracility, Australopithecus garhi (“surprise” in the Afar language). The teeth are slightly larger than those of afarensis (See Figure 4). In overall body shape, however A. garhi was far more modern, with the ratio of upper arm length to upper leg length much closer to those of later hominins (Asfaw et al. 1999). Slightly later in time, from around 2.2 million years ago, came Australopithecus boisei. This form was originally found in 1959 by Mary Leakey, in the Olduvai Gorge area near the Serengeti Plains, in Tanzania, where she and her husband, Louis, had been digging since 1951. This is known as a hyper-robust australopithecine. Comparisons with A. aethiopicus strongly suggest that the facial architecture of A. boisei is very similar, yet larger in size. This has led researchers to surmise that there is an ancestor/descendent link between the two species. A. boisei has a slightly larger cranial capacity of 510 cubic centimeters but otherwise retains all of the characteristics of A. aethiopicus, including the sagittal crest, the scooped-out facial appearance and wide, flaring cheek bones (see Figure 5). It has been hypothesized that these forms subsisted primarily on a vegetative diet consisting of hard nuts, roots and berries because the rear teeth were much larger than the front ones and considerably larger than those ofA. afarensis. This eventually led to the nickname “nutcracker man.” This hypothesis has recently been challenged, however, by research that suggests that the primary diet of this hominin was grasses (Cerling et al. 2011). A. Boiseiis found down to approximately 1.2 million years ago.
Slightly later in time, from around 2.2 million years ago, came Australopithecus boisei. This form was originally found in 1959 by Mary Leakey, in the Olduvai Gorge area near the Serengeti Plains, in Tanzania, where she and her husband, Louis, had been digging since 1951. This is known as a hyper-robust australopithecine. Comparisons with A. aethiopicus strongly suggest that the facial architecture of A. boisei is very similar, yet larger in size. This has led researchers to surmise that there is an ancestor/descendent link between the two species. A. boisei has a slightly larger cranial capacity of 510 cubic centimeters but otherwise retains all of the characteristics of A. aethiopicus, including the sagittal crest, the scooped-out facial appearance and wide, flaring cheek bones (see Figure 5). It has been hypothesized that these forms subsisted primarily on a vegetative diet consisting of hard nuts, roots and berries because the rear teeth were much larger than the front ones and considerably larger than those ofA. afarensis. This eventually led to the nickname “nutcracker man.” This hypothesis has recently been challenged, however, by research that suggests that the primary diet of this hominin was grasses (Cerling et al. 2011). A. Boiseiis found down to approximately 1.2 million years ago. As noted in the last post, the discovery of the Taung child in South Africa fueled interest that other hominins could be found as well. With the help of noted anatomist Robert Broom, and anthropologist John T. Robinson, Raymond Dart excavated several other sites in the South African cave system, Makapansgat, Swartkrans, Sterkfontein and Kromdraai, all of which yielded australopithecine remains of both an early gracile form and later robust form. The earlier form, dated to between 3.0 and 2.0 million years ago and to which the Taung find belonged, was calledAustralopithecus africanus (see Figure 6). Exact dating for these cave sites is hampered by the fact that the cave openings are vertical and it is not clear how the hominins got there. Unlike the east African forms, A. africanus was a very lightly built form, with no crests of any kind. The overall cranial capacity of the finds is between 430 and 520 cubic centimeters, slightly larger than that of A. afarensis and comparable to the east African forms. Like the east African forms, the teeth of this form were larger in the back and smaller in the front than A. afarensis suggesting to some an herbivorous diet.
As noted in the last post, the discovery of the Taung child in South Africa fueled interest that other hominins could be found as well. With the help of noted anatomist Robert Broom, and anthropologist John T. Robinson, Raymond Dart excavated several other sites in the South African cave system, Makapansgat, Swartkrans, Sterkfontein and Kromdraai, all of which yielded australopithecine remains of both an early gracile form and later robust form. The earlier form, dated to between 3.0 and 2.0 million years ago and to which the Taung find belonged, was calledAustralopithecus africanus (see Figure 6). Exact dating for these cave sites is hampered by the fact that the cave openings are vertical and it is not clear how the hominins got there. Unlike the east African forms, A. africanus was a very lightly built form, with no crests of any kind. The overall cranial capacity of the finds is between 430 and 520 cubic centimeters, slightly larger than that of A. afarensis and comparable to the east African forms. Like the east African forms, the teeth of this form were larger in the back and smaller in the front than A. afarensis suggesting to some an herbivorous diet. Slightly more recent in time is A. robustus. This is a scaled-up version of A. africanus and continues the trends seen in that form (See Figure 7). Although having a generally similar facial structure as A. africanus, A. robustus also has crests like the eastern African forms, suggesting a similar dietary adaptation. There is also a continuation of the trend toward larger back teeth and smaller front teeth to accommodate this. While A. robustus and A. boisei are both considered “robust” australopithecines, the considerable differences in their facial architectures suggest to most researchers different evolutionary trajectories.
Slightly more recent in time is A. robustus. This is a scaled-up version of A. africanus and continues the trends seen in that form (See Figure 7). Although having a generally similar facial structure as A. africanus, A. robustus also has crests like the eastern African forms, suggesting a similar dietary adaptation. There is also a continuation of the trend toward larger back teeth and smaller front teeth to accommodate this. While A. robustus and A. boisei are both considered “robust” australopithecines, the considerable differences in their facial architectures suggest to most researchers different evolutionary trajectories. A recent discovery has been made by Lee Berger and colleagues (Berger et al. 2010) at the site of Malapa in South Africa, of yet another form of Australopithecus, which cannot be accommodated into the sequence of A. africanus and A. robustus, and is different enough from each of them to warrant its own species designation: A. sediba. This form is characterized by having a small cranium with no crests and a flat face, in contrast with all other known australopithecines (See Figure 8). Berger suggests that this form is descended from A. africanus and existed parallel to A. robustus. It has also been suggested that there are post-cranial elements in the hip and leg bones that align it with later Homo, although, at this point, the evidence for this position is scant.
A recent discovery has been made by Lee Berger and colleagues (Berger et al. 2010) at the site of Malapa in South Africa, of yet another form of Australopithecus, which cannot be accommodated into the sequence of A. africanus and A. robustus, and is different enough from each of them to warrant its own species designation: A. sediba. This form is characterized by having a small cranium with no crests and a flat face, in contrast with all other known australopithecines (See Figure 8). Berger suggests that this form is descended from A. africanus and existed parallel to A. robustus. It has also been suggested that there are post-cranial elements in the hip and leg bones that align it with later Homo, although, at this point, the evidence for this position is scant. The other find of note was a discovery in northern Kenya, in Lomekwi of a mostly complete but crushed cranium and some facial bones of an individual that shows considerable facial flatness and is lacking in any of the specializations present in the robust australopithecines in terms of dietary adaptations (See Figure 9). It was hence called Kenyanthropus platyops (flat-faced man from Kenya). Dated to approximately 3.5 million years ago, little is known about this find. Its cranial capacity is between that of A. afarensis and A. africanus, at around 400 cubic centimeters (Leakey, et al. 2001). Due to the condition of the cranium, it is difficult to draw any inferences about where this find fits in the phylogeny of the early hominins, and some authors are relegating it to a regional variant of A. afarensis (White 2003). Michael Balter, in fact, recently asked the question
The other find of note was a discovery in northern Kenya, in Lomekwi of a mostly complete but crushed cranium and some facial bones of an individual that shows considerable facial flatness and is lacking in any of the specializations present in the robust australopithecines in terms of dietary adaptations (See Figure 9). It was hence called Kenyanthropus platyops (flat-faced man from Kenya). Dated to approximately 3.5 million years ago, little is known about this find. Its cranial capacity is between that of A. afarensis and A. africanus, at around 400 cubic centimeters (Leakey, et al. 2001). Due to the condition of the cranium, it is difficult to draw any inferences about where this find fits in the phylogeny of the early hominins, and some authors are relegating it to a regional variant of A. afarensis (White 2003). Michael Balter, in fact, recently asked the question 

 In 1961, Louis Leakey’s team discovered the parts of a skull, and some hand and foot bones at Olduvai Gorge. Two years earlier, workers had found the spectacular robust Australopithecus boisei skull, OH5, at this same location. These remains, he concluded were different from the known samples of Australopithecus. The brain size of this individual, OH 7, was 630 cubic centimeters (cc), fully a hundred ccs larger than the just discovered, Au. boisei specimen (L. Leakey, 1961). Not only that but in comparison to the australopithecines, the crania were more lightly built, rounder and slightly larger. The back teeth, which were very large in the robust australopithecines, were smaller and more closely approximated our own dentition. The face was, overall, slightly flatter although it still had the “scooped” appearance of the gracile australopithecines (see sidebar). One such skull was OH 24 (short for Olduvai Hominin 24) (Figure 2), which demonstrates the evolved morphology. This cranium was estimated to have a cranial capacity of slightly less than 600 cc, still far larger than any australopithecine species.
In 1961, Louis Leakey’s team discovered the parts of a skull, and some hand and foot bones at Olduvai Gorge. Two years earlier, workers had found the spectacular robust Australopithecus boisei skull, OH5, at this same location. These remains, he concluded were different from the known samples of Australopithecus. The brain size of this individual, OH 7, was 630 cubic centimeters (cc), fully a hundred ccs larger than the just discovered, Au. boisei specimen (L. Leakey, 1961). Not only that but in comparison to the australopithecines, the crania were more lightly built, rounder and slightly larger. The back teeth, which were very large in the robust australopithecines, were smaller and more closely approximated our own dentition. The face was, overall, slightly flatter although it still had the “scooped” appearance of the gracile australopithecines (see sidebar). One such skull was OH 24 (short for Olduvai Hominin 24) (Figure 2), which demonstrates the evolved morphology. This cranium was estimated to have a cranial capacity of slightly less than 600 cc, still far larger than any australopithecine species. After a few years working at Omo, in the early 1970s he took an expedition team to the Koobi Fora region of Lake Rudolf (now Lake Turkana, See Figure 3, a map of site locations in East Africa) and, in 1972, unearthed one of the most famous of all hominin fossils, the KNM-ER 1470 skull (Figure 4) (Day, Leakey, Walker, & Wood, 1975; R.E. Leakey, 1973). Although he initially dated the find to around 2.9 million years old by palaeomagnetism, discrepancies arose in the form of faunal correlations. The pigs that were found in the same level as ER 1470 were securely dated to around 1.8-1.9 million years at other sites. The level was subsequently re-dated by Potassium-Argon and found to be 1.9 million years old, making it coeval with the robust australopithecines of the region (Johanson & Edey, 1981). This has become the settled date of this fossil, relying not just on biostratigraphy (see sidebar) but radiometric dating.
After a few years working at Omo, in the early 1970s he took an expedition team to the Koobi Fora region of Lake Rudolf (now Lake Turkana, See Figure 3, a map of site locations in East Africa) and, in 1972, unearthed one of the most famous of all hominin fossils, the KNM-ER 1470 skull (Figure 4) (Day, Leakey, Walker, & Wood, 1975; R.E. Leakey, 1973). Although he initially dated the find to around 2.9 million years old by palaeomagnetism, discrepancies arose in the form of faunal correlations. The pigs that were found in the same level as ER 1470 were securely dated to around 1.8-1.9 million years at other sites. The level was subsequently re-dated by Potassium-Argon and found to be 1.9 million years old, making it coeval with the robust australopithecines of the region (Johanson & Edey, 1981). This has become the settled date of this fossil, relying not just on biostratigraphy (see sidebar) but radiometric dating. The initial problem this skull created was that it presented very different morphology from any of the Olduvai specimens, having a much larger supraorbital region and larger, inflated cheeks. Originally subsumed within Homo habilis, the discoverers felt that the difference in size between the Olduvai hominins and the larger fossils from Koobi Fora could not be accommodated within one species. The new specimen had a rounder cranium, flatter face and cranial capacity between 700 and 750 cc., a full half again as much as the largest australopithecines and between 50 and 100 cc greater than the Olduvai Homo habilis sample.
The initial problem this skull created was that it presented very different morphology from any of the Olduvai specimens, having a much larger supraorbital region and larger, inflated cheeks. Originally subsumed within Homo habilis, the discoverers felt that the difference in size between the Olduvai hominins and the larger fossils from Koobi Fora could not be accommodated within one species. The new specimen had a rounder cranium, flatter face and cranial capacity between 700 and 750 cc., a full half again as much as the largest australopithecines and between 50 and 100 cc greater than the Olduvai Homo habilis sample. In addition to the large Homo crania found at East Rudolf, a smaller form was found that was similar in size to the Olduvai Homo sample with the same characteristics as the larger East Rudolf sample. One, in particular, KNM-ER 1813 (Figure 5), had a cranial capacity similar to that of the largest australopithecines, around 510 cc. It has been suggested alternately that the large size differences between the larger and smaller forms at this site represent sexual dimorphism (the difference in size between males and females of the same species) (Rightmire, 1993) or that they represent the presence of H. rudolfensis and H. habilis at the same location (Schrenk, et al., 2007).
In addition to the large Homo crania found at East Rudolf, a smaller form was found that was similar in size to the Olduvai Homo sample with the same characteristics as the larger East Rudolf sample. One, in particular, KNM-ER 1813 (Figure 5), had a cranial capacity similar to that of the largest australopithecines, around 510 cc. It has been suggested alternately that the large size differences between the larger and smaller forms at this site represent sexual dimorphism (the difference in size between males and females of the same species) (Rightmire, 1993) or that they represent the presence of H. rudolfensis and H. habilis at the same location (Schrenk, et al., 2007).

 So far, we have focused on early Homoremains in eastern Africa. Now we can turn our attention to the rest of the continent. In 1953, J.T. Robinson discovered skull fragments in an overhang at Swartkrans cave in South Africa, (the site of many australopithecine discoveries) that, when assembled (SK 847, Figure 2) some years later, demonstrated earlyHomo affinities. This find established the range of early Homo to at least 2500 miles, from East Africa down to the southern end of the continent. (Curnoe, 2010). It is thought that this cranium dates to between 1.8 and 1.5 million years ago but the fossil was not found on a habitation floor, the age remains uncertain.
So far, we have focused on early Homoremains in eastern Africa. Now we can turn our attention to the rest of the continent. In 1953, J.T. Robinson discovered skull fragments in an overhang at Swartkrans cave in South Africa, (the site of many australopithecine discoveries) that, when assembled (SK 847, Figure 2) some years later, demonstrated earlyHomo affinities. This find established the range of early Homo to at least 2500 miles, from East Africa down to the southern end of the continent. (Curnoe, 2010). It is thought that this cranium dates to between 1.8 and 1.5 million years ago but the fossil was not found on a habitation floor, the age remains uncertain. The first cache of these specimens were discovered at Koobi Fora near the shores of Kenya’s Lake Turkana (on the northern border of Kenya) in the 1970s, the most notable of which were KNM-ER 3733 and KNM-ER 3883 (Figure 3).
The first cache of these specimens were discovered at Koobi Fora near the shores of Kenya’s Lake Turkana (on the northern border of Kenya) in the 1970s, the most notable of which were KNM-ER 3733 and KNM-ER 3883 (Figure 3). When we are born, our arm and leg bones are a mix of bone and cartilage, similar to what we have in our nose. As we grow, the cartilage is replaced by bone, which spreads from the middle to the ends of each bone, with the ends “fusing” just before adulthood. This is why it is very easy for us to break bones in childhood. Using the timing of these changes, we can determine how old an individual was at the time of death. In the Nariokotome skeleton, the ends of the bones of the arms and legs (long bones) were not yet fused, indicating that this individual had not reached adult status. The discoverers (Brown, Harris, Leakey, & Walker, 1985) estimated that his adult height would have been nearly six feet. This is significantly greater than the height of any known australopithecine or Homo habilis individual.
When we are born, our arm and leg bones are a mix of bone and cartilage, similar to what we have in our nose. As we grow, the cartilage is replaced by bone, which spreads from the middle to the ends of each bone, with the ends “fusing” just before adulthood. This is why it is very easy for us to break bones in childhood. Using the timing of these changes, we can determine how old an individual was at the time of death. In the Nariokotome skeleton, the ends of the bones of the arms and legs (long bones) were not yet fused, indicating that this individual had not reached adult status. The discoverers (Brown, Harris, Leakey, & Walker, 1985) estimated that his adult height would have been nearly six feet. This is significantly greater than the height of any known australopithecine or Homo habilis individual. Just slightly after H. ergaster appeared on the landscape, a different style of stone tool technology, the
Just slightly after H. ergaster appeared on the landscape, a different style of stone tool technology, the 
 Beginning in the summer of 1999, however, the remains of six individuals—including two remarkable partial skulls, or crania—were recovered from a hilly location at the site of Dmanisi in the Republic of Georgia (see map, below). One surprising characteristic of the skulls was their primitive appearance (Figure 2). Their small cranial size of 800cc put them squarely within the early Homo range (modern humans skulls range between 1350 and 1500 cubic centimeters). Also notable was that from the back, the widest part of the head was quite low, just above the ear. In modern humans, the widest part is three-quarters of the way up from the cranial base. These characteristics, plus the shape of the eye sockets and cheeks strongly suggested an affinity toHomo ergaster (Gabunia et al., 2000).
Beginning in the summer of 1999, however, the remains of six individuals—including two remarkable partial skulls, or crania—were recovered from a hilly location at the site of Dmanisi in the Republic of Georgia (see map, below). One surprising characteristic of the skulls was their primitive appearance (Figure 2). Their small cranial size of 800cc put them squarely within the early Homo range (modern humans skulls range between 1350 and 1500 cubic centimeters). Also notable was that from the back, the widest part of the head was quite low, just above the ear. In modern humans, the widest part is three-quarters of the way up from the cranial base. These characteristics, plus the shape of the eye sockets and cheeks strongly suggested an affinity toHomo ergaster (Gabunia et al., 2000).

 Spain
Spain Evidence for the earliest settlements in Italy has also been discovered in the last decade. At the site of Pirro Nord, a small collection of flakes and scrapers was found at a
Evidence for the earliest settlements in Italy has also been discovered in the last decade. At the site of Pirro Nord, a small collection of flakes and scrapers was found at a Eugene Dubois
It was 1890 and Eugene Dubois was tired. He had searched everywhere in Sumatra for the human ancestors that were supposed to be there—at least according to the theories of his mentor, famed German naturalist Ernst Haeckel. Instead, he had found only heat and malaria.
13 years before, in 1877, Dubois had arrived in Amsterdam to study medicine, but always harboring a desire to study the ancestry of modern humans. So, after four years at the University there, he accepted an invitation to go to the University of Utrecht to study comparative anatomy and devote himself to the latest thinking about the origins of the human species. It was during his time at Utrecht (from 1881 to 1887) that Dubois became enamored of Haeckel’s views on human origins, which differed from those of Darwin. While Darwin argued that humans had evolved in Africa, the region in which our closest living relatives—the chimpanzees and gorillas—still live, Haeckel believed that the origins of humanity lay in East Asia. This was so, he believed, because of his own observations of gibbons that walk bipedally when on the ground.
Haeckel also believed that there had once been a large landmass called Lemuria between the continents of Africa and Asia. In his view, Lemuria had since become submerged, leaving the modern islands of Madagascar and the East Indies as its only remains. The idea of submerged continents was not unusual for the late 19th-century, as people struggled to understand the character of biological diversity present in the world and why there were such striking similarities between animals that were geographically dispersed. The geographical distribution of marsupial fossils in South America and Australia is an example of this sort of problem, and one that was not solved until the second half of the 20th century when continental drift reconstructions suggested that ancient marsupials had used Antarctica as a conduit between the other two continents. Not only did such theories make sense of modern distributions, they were confirmed with later discoveries of marsupial fossils in Antarctica.
In any case, in 1888 Dubois joined the army and set out for the Dutch East Indies to pursue his ideas. For the next two years, he would comb Sumatra attempting to locate the hominin remains that Haeckel promised would be there. In hindsight, what Dubois was attempting was something that had never been done before: discovery of hominin material through the tools of archaeological excavation. Up to this point, all of the human fossils had been found on the surface, eroding out of the side of a bank, or as a result of farming. It had not occurred to anyone to go looking for human ancestors.
Now, with his supply of prison workers dwindling due to desertion and fever, he had almost run out of options and was on the verge of failure. Using almost all of his remaining resources, he decided to abandon his excavations on Sumatra and turn to the nearby island of Java. Emboldened by the fact that early modern human fossils had been discovered there (at Wadjak), he arrived and settled in at Trinil, on the banks of the Solo River, in 1890.

Figure 1: Dubois’ Pithecanthropus erectus
The very next year, Dubois’ long-standing efforts were finally rewarded, first with the discovery of a skullcap (calvaria) of a hominin cranium, and then with an intact femur (Figure 1). Judging by what he knew of cranial anatomy, Dubois estimated that the skull would have been approximately 900 cubic centimeters (cc) in volume, placing it below even the lowest threshold of modern humans. Further, he noticed that it was not like modern humans in shape, being too long and low. He concluded that it showed “evidence of a form intermediate between man and the anthropoid apes” (Dubois, 1896). Dubois envisioned a sequence of forms in which the gibbon gave rise to a form of chimpanzee called Anthropopithecus sivalensis, which then gave rise to the form represented by the Trinil remains, after which Homo sapiens arose (Turner, 1895).
Dubois spent the next twenty years on the road with his find, trying to drum up support for its place in human prehistory. As with Raymond Dart’s discovery of the first australopithecine thirty-three years later, Dubois did not receive a warm reception. Most critics simply said that he had gotten it wrong and that the femur did not belong to the same individual as the obviously-primitive skull cap. Some of the criticism Dubois suffered could have been mitigated had he been more open to sharing the Trinil materials; but, instead, he allowed very little access to the bones, so that very few people knew exactly what they looked like. Adding to Dubois’s credibility problems was the 1911 “discovery” of Piltdown. This intentional hoax turned the paleoanthropology world on its head for forty years, sending researchers down innumerable rabbit holes. As I noted in a previous post, the Piltdown remains made all of the other hominin finds appear too “ape-like” to be on the road to humanity and informed many opinions about finds such as those from Trinil.
On the other hand, some critics of Dubois’ new hominin claim were vicious, and questioned both his academic abilities and his judgment (Shipman & Storm, 2002)—in addition to the interpretation of the find itself. It was in reference to Dubois’ work that the term “Missing Link” was first used with reference to a particular human fossil, originating with Charles Lyell (1863) and describing palaeontological gaps. And ironically, it was in one of the most stinging criticisms of Dubois’ work that the name that would eventually stick was first used: “Homo erectus.” Eventually, many other finds in the same general area and across Southeast Asia demonstrated that what Dubois had found was a real, previously-unknown hominin form, and the first to colonize the Asian continent and the islands leading off towards Oceania.
Homo erectus across South East Asia

Figure 2: Sangiran 17
Sangiran
The earliest point at which Homo erectus appears to have begun to colonize the greater East Asian region is around 1.8 million years ago, represented first by the partial child’s skull found at the site of Modjokerto, and then, at around 1.66 million years ago, at the site of Sangiran, in Trinil, where Dubois had made his landmark discovery. This site was rich, yielding the remains of many crania, perhaps best represented by Sangiran 17 (Figure 2), an almost complete skull.
The material from the Sangiran site is very diverse morphologically, with some crania having capacities of as little as 700 to 800 cc, and other, larger heads with volumes in the range of 1000 cc. As with the late Homo ergaster finds from Africa, the remains from Sangiran yielded crania that were still widest at their bases, possessing large brow ridges. Some have thick cranial bones and are very robust (Sangiran 4), while others are very gracile (Sangiran 31). What this variation means is not clear, but most workers believe it represents a very diverse diachronic population (that is, one group living and moving around over a long period) rather than separate species inhabiting the area. The Sangiran site yielded fossil material in an almost continuous succession from approximately 1.66 million years ago to less than 800,000 years ago.
Because the area of the excavations—the Sangiran Dome—is a volcanic deposit, the layers have been securely dated by the 40Ar/39Ar method, although questions remain about the historical sequence and distribution of other animals that lived there through the ages (its faunal succession). The problem is that many of the fossils were not found in context, and relating them directly to the stratigraphy is tenuous. Despite this, most workers are comfortable with the earliest hominins in the region being at least 1.5 million years old.
One of the things hampering workers in this region is the comparative paucity of recovered stone tools. Those that have been found suggest a technological stage similar to the late Oldowan design, equivalent to that being created by the Homo ergaster populations inhabiting the area of Dmanisi and East Africa. Unfortunately, none of the tools have been associated with the hominins directly so it is not exactly clear who made them.
 Figure 3: Sambungmacan 3
Figure 3: Sambungmacan 3
Sambungmacan
Another major find from the area where Dubois brought Homo erectus to light is the cranium from the site of Sambungmachan. This skull was reportedly found in 1977 but was then illegally sold to the antiquities market, where is spent considerable time in different collections before being “rediscovered” in 1998—in a New York nature curio shop called Maxilla and Mandible, Inc. (Delson et al., 2001). This was an almost-complete calvaria (Figure 3), with only part of the base missing. It is equivalent in size to the fossils from Sangiran, with a cranial capacity of around 1000 cc. It has a large brow ridge extending all of the way across the top of the eyes, a long, low cranium with a sloping forehead and a maximum width near the cranial base—all features that are also characteristic of the late African H. ergaster and Sangiran crania. Although we will never know exactly how old this cranium is, its morphology is consistent with that of the material from Sangiran.
 Figure 4: Ngandong 6
Figure 4: Ngandong 6
Ngandong
Later in time, but also located on the Solo River, is the site of Ngandong, excavated by Oppenoorth in the early 1930s. At this site, fourteen calvaria have been discovered, all of which show advanced Homo erectus characteristics: long and low in shape, with thick-bones and a distinctive brow-ridge. (Figure 4). As with the other Indonesian finds, dating the Ngandong material has been problematic. The deposits at the site were originally thought to be around 100,000 years old, but this interpretation was turned on its head in 1996, when Swisher and colleagues claimed that the deposits were no older than between 27,000 and 53,000 years old (Swisher et al., 1996). These age estimations were made on the associated fauna, however, and as Rainer Grün and the late Alan Thorne pointed out, the faunal material does not match the skulls either in color or in texture and is likely not from the same time. Recently, Swisher and colleagues revisited the dating of the site and derived internally-consistent dates of at least 143,000 years before the present (Indriati et al., 2011). As with the Trinil remains, however, there are no associated stone tools.
Homo erectus in China
The Chinese Homo erectus material is very widely scattered and working in the region has presented many difficulties for researchers in terms of transport, language barriers and funding. Consequently, we know less about this region and its previous inhabitants than we do about most other areas of the Old World. Although there are between ten and fifteen sites that have yielded Homo erectus material, I will only touch on the most important ones.

Figure 5: Lantian
Lantian
In the early 1960s, a cranium and mandible were found in the cave of Lantian, Shaanxi province, whose characteristics matched other remains from China designated as Homo erectus. Paleomagnetic dating has yielded a date no earlier than 1.15 million years ago for the skull, with the consensus being that it is around 800,000 years old. A date of approximately 650,000 years before the present was derived for the mandible. The cranium is heavily encrusted and suffered from postmortem deformation (Figure 5). When reconstituted, it was found to have a capacity of around 780 cc (low for Homo erectus) and the bones on the sides of the head are the thickest yet recorded. At this site some flake tools, mammal remains, and an ash deposit were all recovered, suggesting hunting and control of fire.
 Figure 5: Hexian
Figure 5: Hexian
Hexian
Another almost-complete calvaria was found at Longtandong cave in the province of Hé Xiàn, dated to between 400,000 and 500,000 years ago. This find exemplifies typical Homo erectus in many ways in that it is long and low, with heavy muscle markings toward the base and the rear of the skull (Figure 6). The cranial capacity is around 1000 cc, a third-again greater than that of the Lantian calvaria. Its cranial shape is very similar to those found in Southeast Asia, suggesting that it straddles the Southeast Asian and Chinese boundary.
While both Lantian and Hexian were significant finds, another site in China boasted the single largest collection ofHomo erectus fossils ever found at one site, as well as presenting one of the greatest mysteries in paleoanthropology. In the conclusion of our look at Homo erectus in Asia, we’ll peer into the Zhoukoudian caves and consider how this species fits into the lineage of man.
Zhoukoudian

The Zhoukoudian site yielded an amazing amount of material: twelve total crania of which six were almost complete, as well as a large collection of mandibles and hundreds of cranial fragments (Figure 7). The cranial capacity of these individuals ranges from 915 to 1225 cc, making them larger than the majority of the Southeast Asian individuals. But nearly as important as the fossils themselves, researchers found numerous stone “cleavers” and four layers of ash, reflecting the use of fire. All in all, there seems to have been a continuous occupation of the cave from around 600,000 to 200,000 years ago, spanning thirteen layers and some forty meters of deposits. Along with the crude implements, hundreds of mammal species are represented by remains in the cave, many of the bones having distinctive cut marks on them. This suggests that the occupants of this cave could (and did) hunt many different species for food.
Homo erectus and the Hand Axe
While the archaeological record of tools for the Southeast Asian Homo erectus sample is extremely thin, some Chinese sites have provided artifacts. (Schick & Zhuan, 1993). Uniformly crude and not as developed as the Acheulean mode tools in East Africa, these bifacially-worked chopping tools have been referred to as “cleavers” by some researchers (figure below). There is considerable variation in size and shape, however, and Lycett has argued that there are regional variations in biface design and construction that can be attributed to the demands of different environments (Lycett, 2008). Interestingly, it is also clear that, over time, these tools became more uniform, resembling the hand axes that are found elsewhere. This suggests two strong possibilities: first, that specific functional necessities were the driving force behind their shape, and that these functional constraints necessitated a similarity in design between groups; or, second, that “technology swapping” between populations was occurring.
Figure 8: Bifaces from Zhoukoudian

Although these “cleavers” appear in numerous places in the Chinese record, there is a sharp break between them and the well-made hand axes of Africa, Southwest Asia and the Indian sub-continent. This break is referred to as the “Movius” Line, named for the researcher who first noticed the pattern. It has always been a source of puzzlement to archaeologists, but recently, some researchers have suggested that this demarcation exists because the Chinese Homo erectus populations possessed a raw material alternative for creating the complex tools they needed: bamboo. Bar-Yosef and colleagues have shown that with less effort and time required to make them, bamboo points can be created that are nearly as strong as and easier to use than stone ones (Bar-Yosef, Eren, Yuan, Cohen, & Li, 2012).
Beyond Homo ergaster: How many species of Mid-Pleistocene Homo?
One of the persistent problems of hominin studies when we get to our own line is attempting to determine how many species coexisted. As we have seen previously, there was even disagreement about how many species were represented in Eastern Africa, with Homo rudolfensis and Homo ergaster.
Historically, many archaeologists have held that Homo erectus was the hominin that colonized the Old World, and that this colonization took place between one and two million years ago. During the 1970s and 1980s, paleoanthropological thought held that Homo habilis followed the reign of the australopithecines, and was in turn followed by Homo erectus. More recently consensus has emerged around the idea that there was further speciation in those populations that ended up in Europe and Asia. Whether one accepts one species (H. erectus) or two (H. erectus and H. ergaster), there is, as Ian Tattersall describes, “a relatively cohesive subset of the family Hominidae.” (Tattersall, 2007)
Indeed, one is struck by the unity in morphological features that make up Homo erectus/Homo ergaster in all parts of the Old World. Rightmire (Rightmire, 1998) has consistently argued that the traits used to differentiate Homo ergaster from Homo erectus are lacking and that it is best to simply use the name Homo erectus to describe all of the middle Pleistocene hominins between about 1.6 million and 300,000 years ago. Antón suggests the following traits that characterize the species as a whole: a cranial vault ranging in volume from approximately 700 to 1200 cc; a low, sloping forehead and sharply angled rear; a large, single brow ridge over the eyes; and sharp muscle markings on base of the vault with very thick cranial bone. (Antón, 2003).
Cranial morphology is remarkably static through time as well (Kidder & Durband, 2004), with little change for hundreds of thousands of years. Differences that are found do not constitute trait polarities (presence or absence of a particular feature), but are differences in overall dimensions within a consistent general type, such that it is reasonable to assign these distinctions to regional genetic drift.
On the other hand, Wood has made the argument that, while it is reasonable to assign the taxon Homo erectus to the material in Asia, it is not reasonable to do so for the African remains: the Asian samples are marked by the presence of traits such as the sagittal keel (a ridge of bone on the top of the head, visible on Figures 1 and 4), the angular torus and thick cranial bone, while these features are not as evident in the African examples. Wood has suggested that the earliest fossils from Africa—originally designated Homo erectus—be called Homo ergaster, instead, a convention I adopted for the previous post in this series. At some point, however, a group or groups left Africa with some of these traits present, and the best explanation for their presence in both the Chinese and South East Asian populations is shared retentions from an ancestral population.
Homo erectus and the Created Order
Figure 9: Zhoukoudian Homo erectus and
Australopithecus africanus

At some point we must consider Homo erectus from another perspective, as well. It has been argued by those opposing evolution in general and human evolution in particular that the australopithecines represent nothing more than aberrant apes (Morris, 2002). While such an argument does not bear up under scrutiny even with regard to australopithecines, itcertainly cannot be used to describe Homo erectus in light of the extensive material we have for this species. Not only is Homo erectus morphologically stable through time and across geographical space, it is also clear that these hominins are considerably more advanced than australopithecines (Figure 9). The crania of Homo erectus are longer, higher and considerably larger in volume; the face is more pulled in and less scooped. These are individuals that are creating complex stone tools, which are economical and efficient. In China, they are also controlling fire, an invention that completely changes how the day may be organized, and also expands the repertoire of what is edible. There is evidence for the controlled use of fire at both Lantian and Zhoukoudian and, considering the evidence at Gesher Benot Ya’aqov, it appears that as Homo erectus moved east, they brought fire with them.
While not completely human, it is clear that Homo erectus were hominins practicing many of the behavioral patterns that we identify as human, and were getting closer to being human.